
|
DIVERSITE
ET COMPLEMENTARITE DES METABOLISMES
|
|---|
Herbier de posidonies en méditerranée
(Bonifacio) : les premiers maillons d'une chaine alimentaire.
Les végétaux chlorophylliens sont des producteurs primaires : ils fabriquent leurs constituants organiques à partir de substances minérales et sont dits pour cela "autotrophes" par opposition aux autres êtres vivants "hétérotrophes".
Pb : Comment cette autotrophie est-elle possible ?
I- La synthèse de matière organique
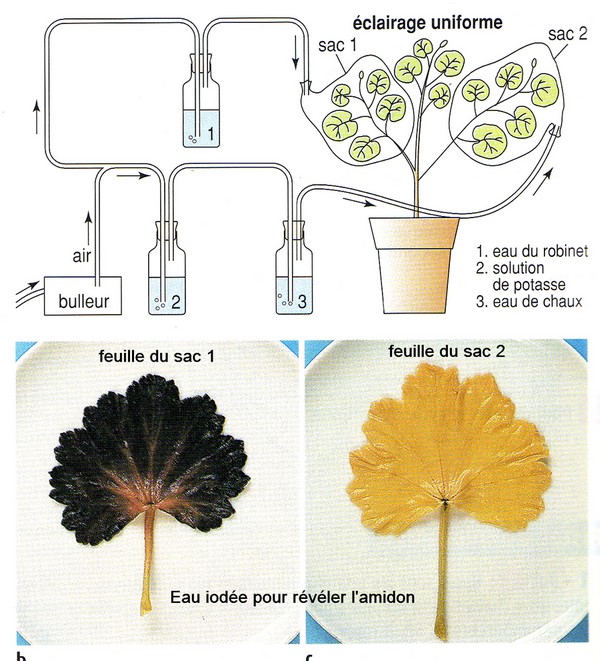
1- Localisation de la photosynthèse dans une plante éclairée
En démo au bureau : feuilles de géranium éclairées 24h avec cache + témoin à l'obscurité

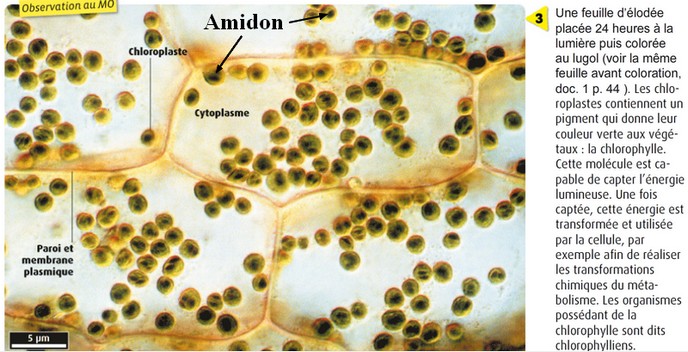

La
synthèse de matière organique est photodépendante
: elle ne se produit qu'à la lumière dans les chloroplastes
des cellules végétales. Cette matière organique est une macromolécule glucidique :
l'amidon, révélé par l'eau iodée.
2- Recherche de la source de carbone
Manuel p178, 179

Documents extraits du manuel de SVT spe Bordas 2002 p178-179
Chez les végétaux chlorophylliens, le
carbone utilisé pour synthétiser l'amidon est issu d'une molécule minérale oxydée : le CO2
atmosphérique ou l'ion hydrogénocarbonate HCO3-
dans l'eau.
Les êtres vivants capables de produire de la matière
organique en procédant à la réduction d'une matière minérale
(CO2
ou HCO3-) sont dits autotrophes
pour le carbone.
Remarque
: l'autotrophie peut également concernée l'azote : un organisme capable
de réduire l'azote minéral (N2 ou NO3) est dit autotrophe pour
l'azote...
Pb : la source d'énergie nécessaire à cette synthèse de matière organique est-elle toujours la lumière ?


| Les végétaux chlorophylliens utilisent l'énergie lumineuse du soleil pour réaliser la synthèse de matière organique : on parle de phototrophie ou plus généralement de photosynthèse. Dans certains écosystèmes particuliers comme au voisinage des sources hydrothermales des grands fonds océaniques, il existe des bactéries autotrophes qui tirent leur énergie de l'oxydation de molécules inorganiques (hydrogène, sulfure d'hydrogène) : on parle alors de chimiotrophie ou plus généralement de chimiosynthèse. |
II- Les échanges gazeux chez un végétal chlorophyllien
1- Mise en évidence des échanges gazeux d'une plante verte aquatique (l'élodée) à l'obscurité puis à la lumière |
|
Remarque 1 : les échanges gazeux de la respiration et de la photosynthèse étant contraires, il y existe un point de compensation pour lequel la plante verte ne produit pas plus de dioxygène par photosynthèse qu'elle n'en consomme pour la respiration.
Remarque 2 : dire que la forêt amazonnienne est le "poumon de la Terre" est une idée fausse car c'est oublier que globalement les plantes respirent autant qu'elles font de la photosynthèse et c'est oublier aussi qu'un bois mort est consommé par des millions de microorganismes qui eux aussi respirent!
Pour aller plus loin : http://planet-terre.ens-lyon.fr/planetterre/XML/db/planetterre/metadata/LOM-oxygene-forets.xml
Equation globale de la photosynthèse : 6 CO2 + 12 H2O |
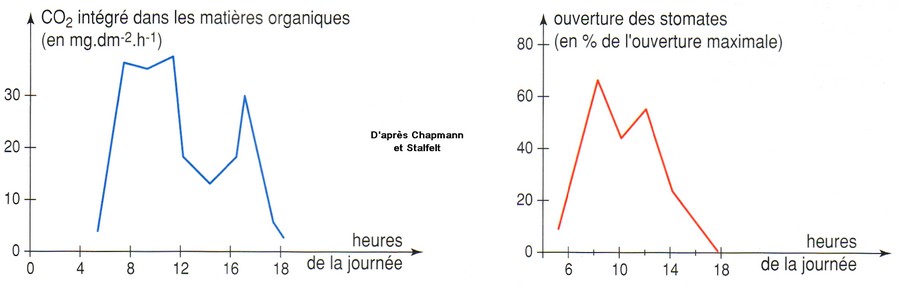
Pb : Comment les cellules chlorophylliennes sont-elles alimentées en CO2 ?
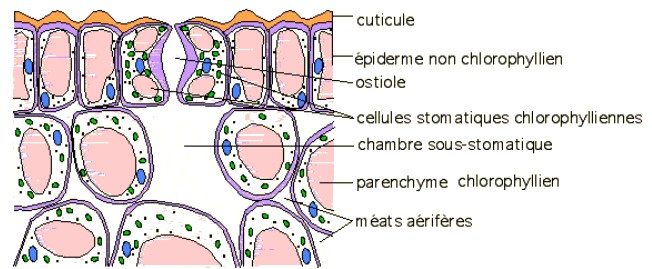
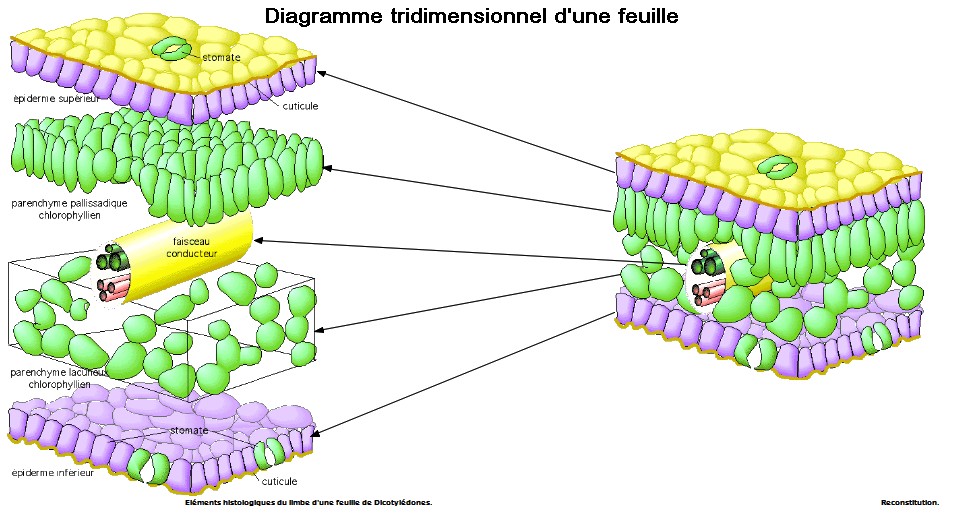
2- Observation microscopique de la face inférieure d'une feuille de polypode


|
|
Remarque : de nombreuses plantes adaptées à la sécheresse (Cactacées, Crassulacées, Euphorbiacées, ...) ont leurs stomates fermés le jour pour limiter les pertes d'eau et ouverts la nuit pour capter le CO2. Leur métabolisme est également différent...
3- Observation microscopique d'une feuille de houx en coupe transversale
TP : réalisation d'une coupe transversale fine dans une feuille de houx :

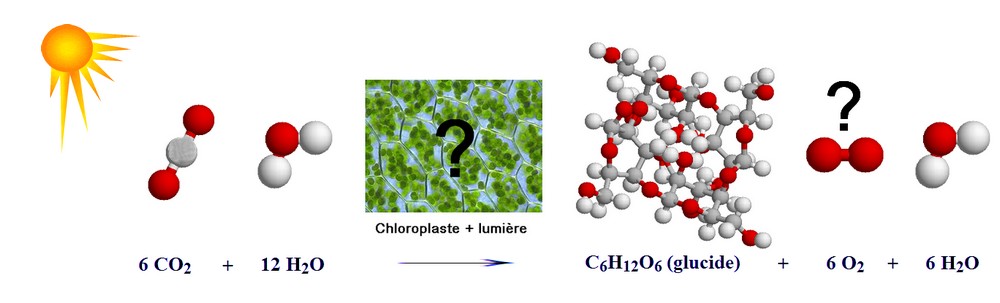
Pb : Comment l'énergie lumineuse est-elle convertie en énergie chimique ? Comment est assurée l'assimilation du CO2 ? Quelle est l'origine du dioxygène libéré ?
III- Les pigments chlorophylliens
1- Localisation des pigments chez les Eucaryotes photosynthétiques (plantes, algues)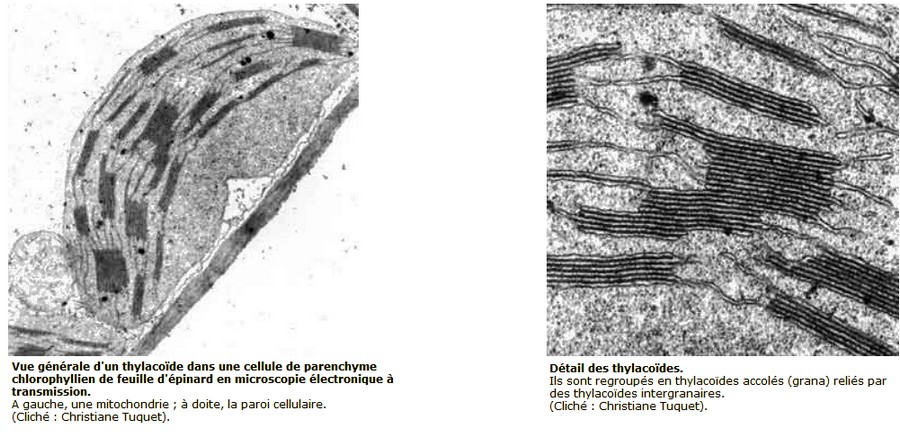
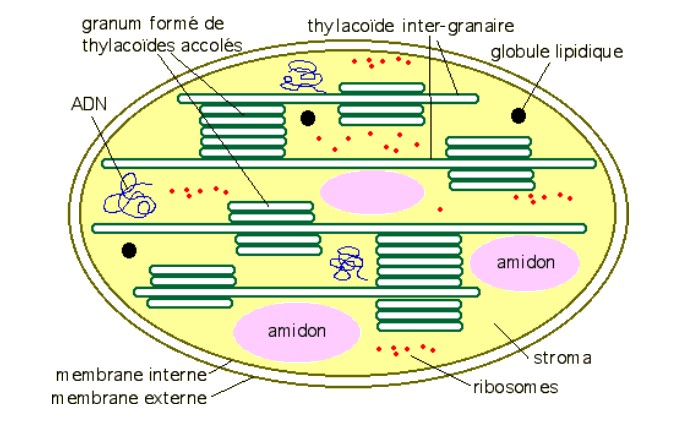
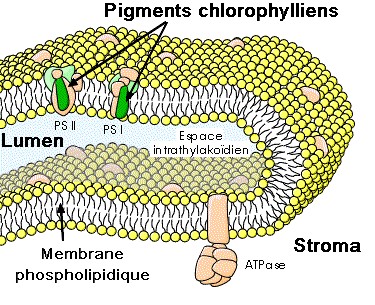
Source : http://www.snv.jussieu.fr/bmedia/Chloroplaste/met.htm
| Les pigments chlorophylliens liposolubles sont enchassés dans la membrane des thylakoïdes qui baignent dans un gel hydraté : le stroma. |
2- Séparation des pigments chlorophylliens par chromatographie
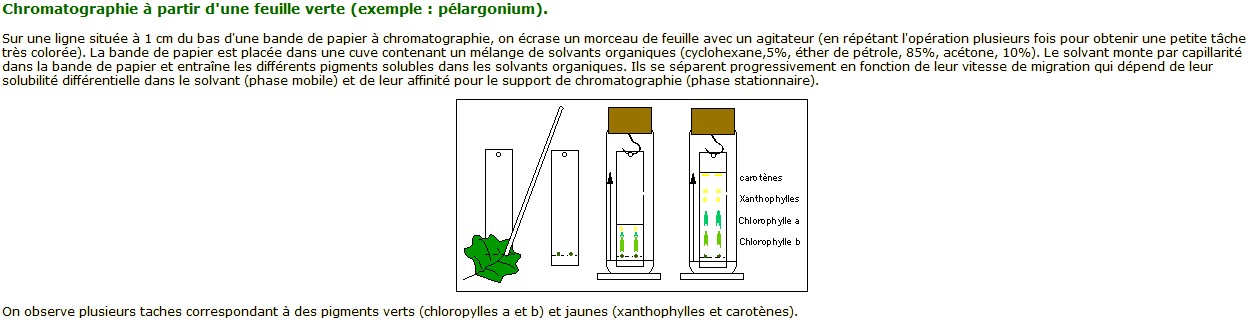
Source : http://www.snv.jussieu.fr/bmedia/Photosynthese/exp22.html
| Par chromatographie, on peut
séparer plusieurs types de pigments dans les feuilles : - Les pigments solubles dans les solvants organiques : chlorophylles a et b (verts), caroténoides (orange). - Les pigments insolubles dans les solvants organiques mais solubles dans l'eau : anthocynanes (bleus). |
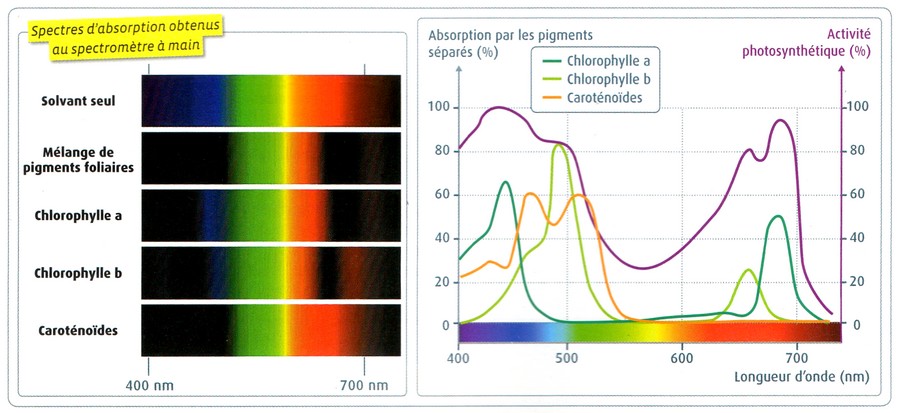
| Les pigments absorbent principalement
la lumière bleue et rouge, ce qui explique la coloration verte des
feuilles. Il y a une bonne correspondance entre le spectre d'absorption des pigments et le spectre d'action photosynthétique : les longueurs d'onde absorbées sont utilisées pour la photosynthèse. |
Pb : quel rôle les pigments jouent-ils dans la photosynthèse ? Quelles sont les étapes de la photosynthèse ?
IV- Les étapes de la photosynthèse
1- Expériences d'Emerson et Arnold (1932)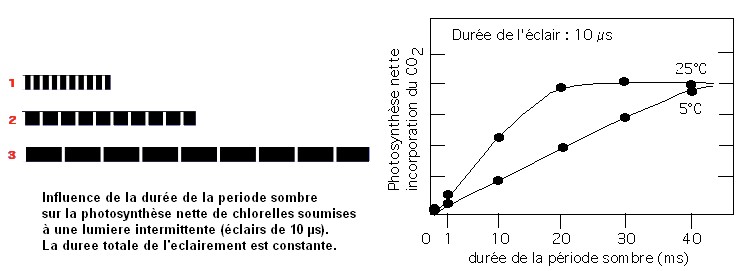
| Dans les conditions de cette expérience (éclairement total bref et saturant), il faut une période sombre importante pour obtenir une photosynthèse maximale. Ceci suggère que des intermédiaires sont formés à la lumière rapidement (de manière quasi insensible à la température) = réactions photochimiques, et qu'ils sont utilisés beaucoup plus lentement par des réactions chimiques (sensibles à la température) = réactions biochimiques d'assimilation du CO2 . |
Pb : en quoi consiste exactement cette phase photochimique ?
2- La phase photochimique : 1ère étape de la photosynthèse
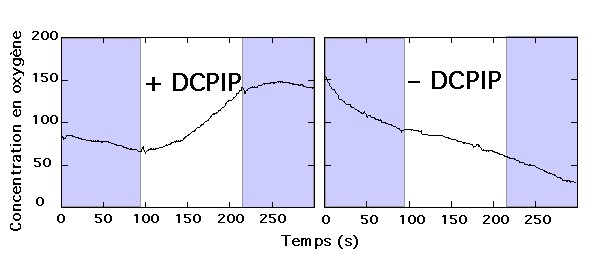
En 1937, Hill réalise son expérience sur une suspension de chloroplastes éclairée et utilise, en absence de CO2, un réducteur artificiel : le ferricyanure de potassium (réactif de Hill)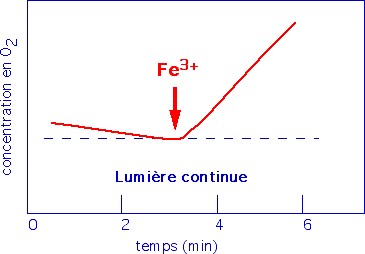 |
En absence de CO2, les chloroplastes sont capables de libérer du
dioxygène, à condition qu'un accepteur d'électron (Fe3+) soit présent
dans le milieu. |
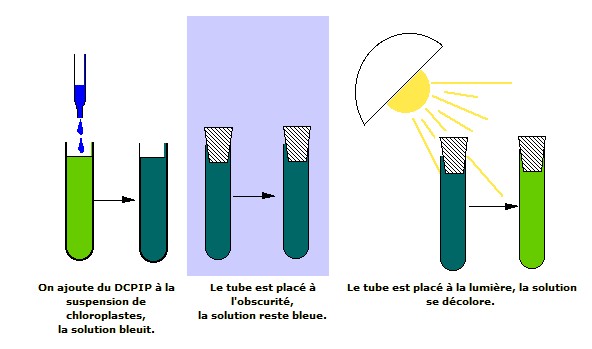 |
|
| Le DCPIP est décoloré, donc réduit à la lumière par la suspension de chloroplastes. | En absence de source carbonée, à la lumière, les chloroplastes isolés produisent un dégagement de dioxygène, parallèlement à la réduction du DCPIP. |
Source : http://www.snv.jussieu.fr/bmedia/Photosynthese/exp44.html
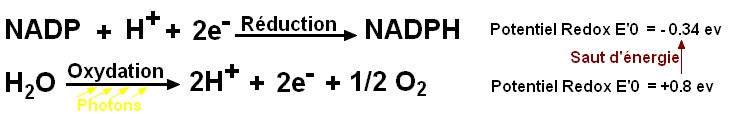
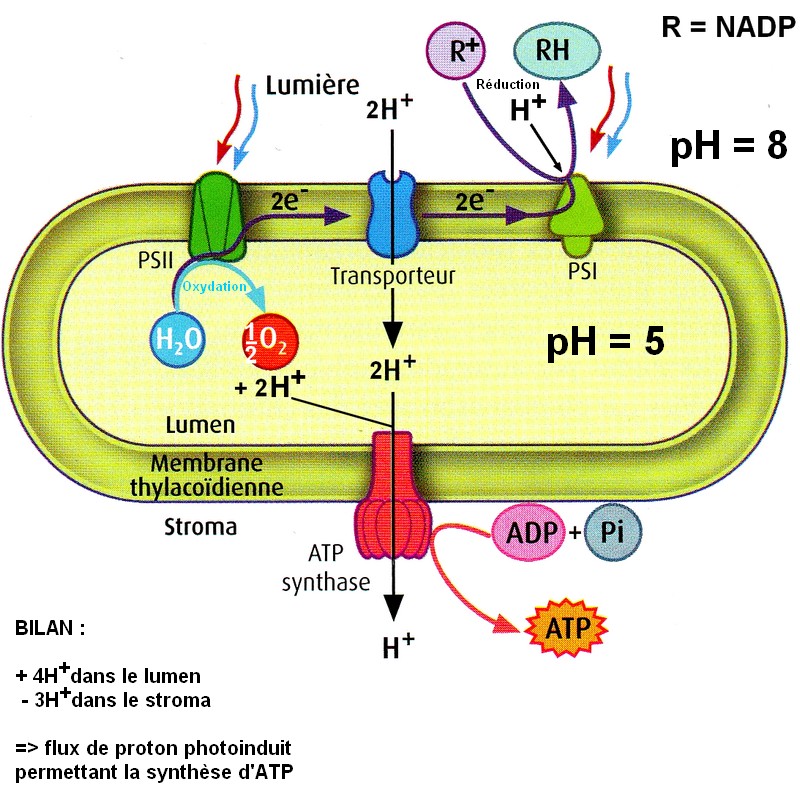
Extrait du manuel de Terminale spe SVT Belin 2012 p21 (modifié)
| la photosynthèse peut donc être séparée en 2 réactions couplées
à un système d'oxydo-réduction. Dans les conditions naturelles de
la photosynthèse, ce rôle d'accepteur d'électron est rempli par le
NADP+ (Nicotinamide Adénie Dinucléotide Phosphate). Le couple NADP+
/ NADPH joue ainsi le rôle d'intermédiaire entre l'oxydation de l'eau
et la réduction du CO2. Il existe en fait une série de couples redox qui transfèrent successivement les électrons jusqu’à un accepteur final, le long d’une chaîne d’oxydo-réductions dans la membrane du thylacoïde. Avec les protons issus de la photooxydation de l'eau, le lumen devient plus acide que le stroma et un flux transmembranaire de H+ permet in fine de synthétiser de l'ATP (Adénosine Triphosphate), molécule portant l'énergie dans ses liaisons P-P. La phase photochimique correspond donc à la synthèse d'un pouvoir réducteur (le NADPH) et d'un pouvoir énergétique (l'ATP). |
Remarque : pour certains auteurs, la photosynthèse au sens strict se limite à la synthèse du pouvoir énergétique et du pouvoir réducteur. Dans ce cours, comme dans beaucoup d'ouvrages, nous inclurons l'assimilation du dioxyde de carbone et la synthèse de molécules carbonées dans la photosynthèse (prise donc au sens large)
Pour aller plus loin (attention, hors programme!) : http://www.snv.jussieu.fr/bmedia/Photosynthese-cours/12-schemaz.htm
3- La phase chimique : 2ème étape de la photosynthèse
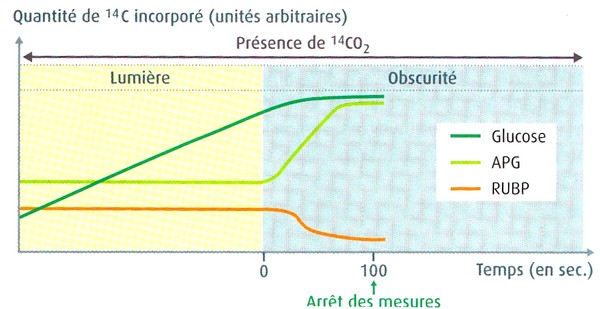
Ce sont les expériences de Calvin (1952) et Bassham et Calvin (1959) qui ont permis de connaître la nature du premier composé carboné formé par la photosynthèse.
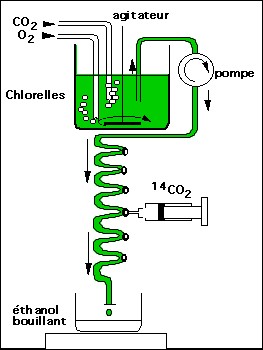
Expérience de
Calvin, protocole expérimental.
En fonction du débit de la pompe, on est capable
très précisément de calculer le temps pendant lequel les chlorelles situées
dans le serpentin ont été en contact avec le CO2 radioactif
avant d'être fixées par le méthanol bouillant.
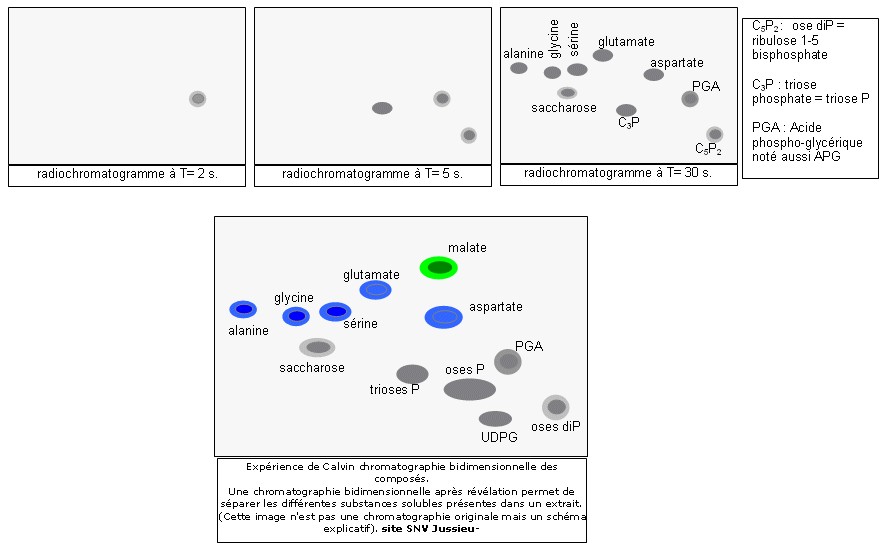
L'incorporation du CO2 se réalise donc de la manière suivante :
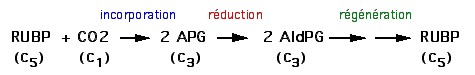
| La phase chimique se déroule dans le stroma des chloroplastes et
se déroule en 3 étapes au cours du cycle
de Calvin : 1ère étape : fixation du CO2 sur un composé organique en 5 carbone, le ribulose biphosphate (RuBP), grâce à une enzyme clé : la Rubisco. le composé en C6 est scindé en deux molécules en C3 : l'APG (Acide phosphoglycolique) 2ème étape : réduction de l'APG en 3-phosphoglycéraldéhyde (G3P) grâce à l'ATP et au NADPH produit à la phase photochimique. 2 G3P (molécule en C3) sur 12 servent à la synthèse du glucose (molécule en C6) 3ème étape : régénération du RuBP à partir du G3P restant grâce à l'ATP produit à la phase photochimique. |
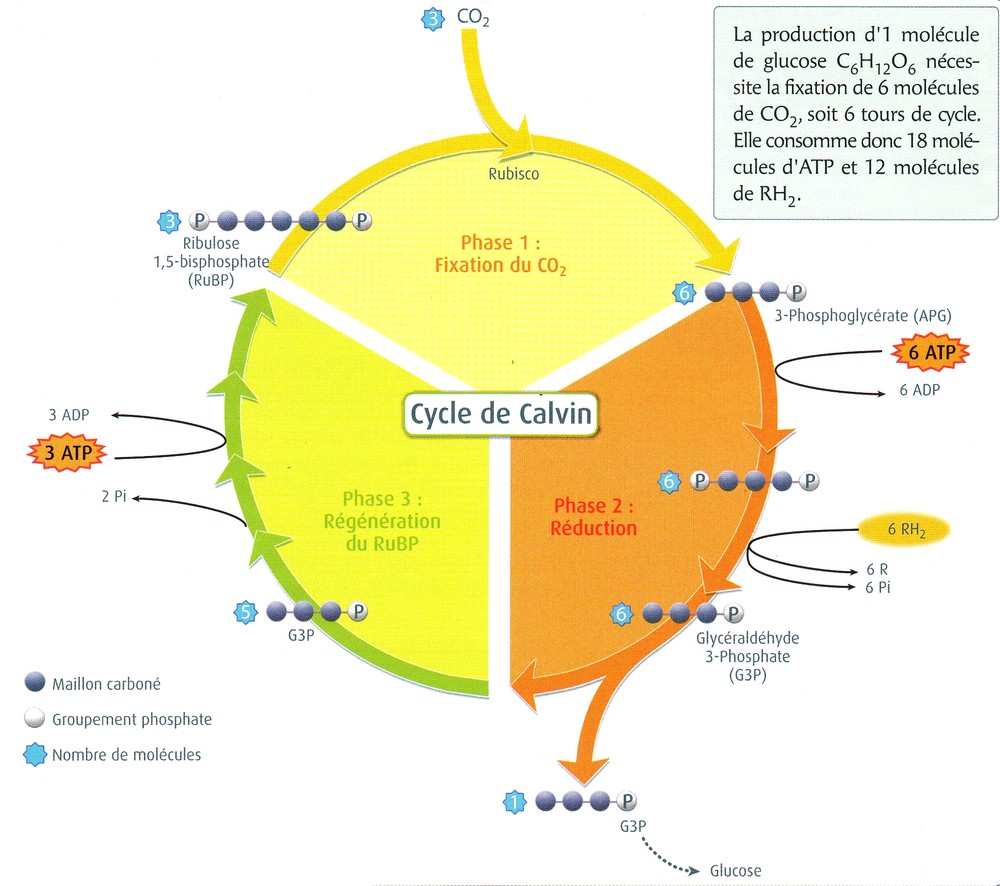
Remarque 1 : la Rubisco est l'enzyme la plus abondante sur Terre
Remarque 2 : cette enzyme est peu spécifique. Elle catalyse aussi bien la fixation du CO2 que celle de l'O2 sur le RuBP!
Remarque 3 : son activité est indirectement dépendante de la lumière. Elle ne fonctionne qu'à la lumière car elle nécessite du Mg2+ qui passe dans le stroma pour compenser alors l'entrée de H+. Enfin, son activité est maximale à pH 8, pH du stroma dans un chloroplaste éclairé...
L'ancienne terminologie "phase sombre" ou "non photodépendante" pour cette phase chimique est donc totalement injustifiée
{kind=link}
{kind=link}