|
La reproduction sexuée permet la transmission des innovations génétiques. L'acquisition d'une innovation dans les futures générations suffit-elle à changer l'espèce ?
Hypothèses : cela doit dépendre sûrement du nombre d'individus concernés, de l'environnement...
I- Un exemple de sélection naturelle : le mélanisme industriel chez la phalène du bouleau
La phalène du bouleau (Biston betularia) est un papillon,
fréquent en Europe du nord, qui vole la nuit et se repose le jour dans
les bouleaux. Cette espèce a été très étudiée
depuis le XIXème siècle car elle présente deux formes
principales : l'une claire (typica) et l'autre sombre ou mélanique
(carbonaria), dont les fréquences ont considérablement varié
dans les populations naturelles au cours des 150 dernières années.
Au milieu du XIXème siècle, la forme typica était largement
majoritaire dans les populations anglaises. En 1848 on a capturé le
premier individu carbonaria dans la région de Manchester.
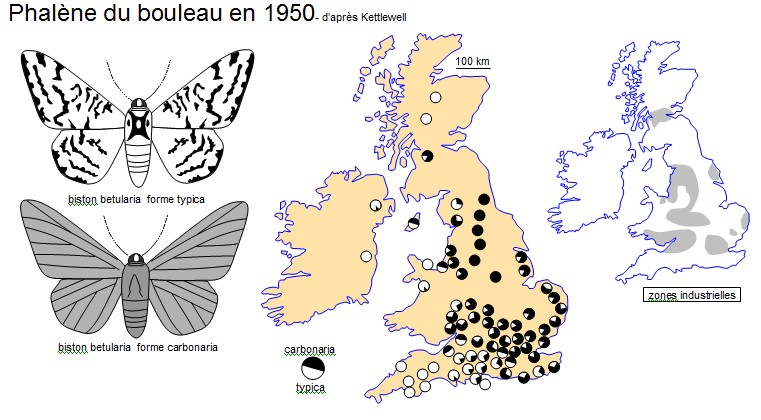
1) En vous appuyant sur le document ci-dessus, quel constat pouvez vous faire ?
La forme "carbonaria" est majoritaire dans les régions industrielles. Il y a peut être un lien entre la couleur du papillon et la couleur de son environnement (la suie noircit les troncs d'arbre) |
A l'époque 2 modèles s'opposaient :
Le modèle lamarckien
L'hypothèse la plus répandue supposait une action dirigée
(non aléatoire) de l'environnement, modifiant un caractère héréditaire
d'un organisme pour mieux adapter celui-ci à son milieu. Le caractère
serait alors transmis sous la forme nouvelle. Cette hypothèse a été
formalisée par LAMARCK (1744-1829). Elle est souvent présentée
sous le terme d'hérédité des caractères acquis.
Le modèle darwinien
Dès 1897, certains chercheurs proposèrent une explication au
mélanisme industriel dans le cadre de la sélection naturelle:
on suggéra que les individus typica, posés sur des bouleaux
clairs couverts de lichens, étaient mieux camouflés des oiseaux
prédateurs que les individus carbonaria. Sur des bouleaux noircis par
les suies et dépourvus de lichens, c'est l'inverse qui se produisait.
Les deux formes seraient alors consommées de façon différente
selon le type de support, la forme mélanique étant avantagée
sur des arbres noircis. Elle se serait ainsi répandue facilement dans
les régions industrialisées, suite à la modification
de la surface des bouleaux par la pollution.
2) Pourquoi le modèle lamarckien n'est-il pas valide, aux vues de nos
connaissances sur le programme génétique ?
|
L' hypothèse sélectionniste fut développée par différents chercheurs et mise à l'épreuve dans les années 1950. On utilisa la méthode des captures-recaptures : un grand nombre de phalènes des deux formes furent marquées ventralement, puis relâchées. Quelques jours après, une nouvelle campagne de captures fut menée, et les individus marqués et non-marqués des deux formes furent dénombrés. Les résultats des expériences de captures-recaptures effectuées dans deux régions proches, mais différentes par leur taux de pollution, réalisées par KETTLEWELL (1955) sont présentés ci-dessous.
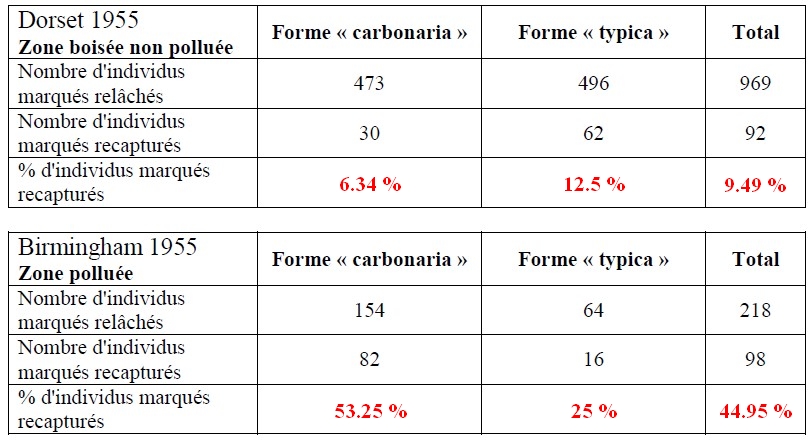
3) Calculez les pourcentages d'individus marqués recapturés dans chaque région. Qu'en déduisez-vous quant aux chances de survie de chaque forme ?
|
Pour vérifier la contrainte sélective portée sur certains papillons dans un environnement donné, KETTLEWELL a exposé à la prédation par les oiseaux le même nombre de papillons des deux formes sur chacun des deux sites et a compté le nombre de papillons capturés par les oiseaux dans chaque cas.

4) Calculez les pourcentages d'individus capturés par les oiseaux dans chaque région puis mettez en relation ces résultats avec ceux obtenus précédemment.
|
La coloration de la phalène est lié à un gène pour lequel il existe 2 allèles : C est l'allèle responsable de la forme "carbonaria" et domine l'allèle c responsable de la forme "typica".
Soit p la fréquence de l'allèle C et q la fréquence de l'allèle c : p+q=1 (100%)
De nombreux travaux ont cherché à modéliser les effets de la sélection naturelle sur la variation des fréquences alléliques, au cours des générations. Le paramètre de base permettant de quantifier l'action de la sélection est appelé valeur sélective ou valeur " adaptative " du phénotype ("Darwinian fitness"). Il est classiquement noté w.
En supposant que les taux de recapture correspondent aux taux de survie, on peut en déduire les valeurs sélectives absolues :
Exemple :
pour 20% de recapture de la forme carbonaria : valeur sélective absolue de l'homozygote dominant = valeur sélective absolue de l'hétérozygote = 0.2
pour 10% de recapture de la forme typica : valeur sélective absolue de l'homozygote récessif = 0.1
Dans la pratique on rapporte les valeurs absolues à la valeur du meilleur génotype (valeur sélective relative) :
w11 = w12 = 0.2/0.2 = 1
w22 = 0.1/0.2 = 0.5
5) Calculez w11, w12 et w22 dans le Dorset puis à Birmingham en 1955
w11 = valeur sélective relative
de l'homozygote C//C |
w12 = valeur sélective relative
de l'hétérozygote C//c |
w22 = valeur sélective relative
de l'homozygote c//c |
|
Dorset |
0.5 |
0.5 |
1 |
Birmingham |
1 |
1 |
0.5 |
6) Utilisez le programme informatique Natural Selection "Kent HOLSINGER" de l'Université du Connecticut pour suivre sur 100 générations l'évolution d'une fréquence allélique p fixée au départ (en 1955) à 0.9 dans le Dorset puis à Birmingham.

7) Au rythme de 1 génération par an, quelle est la fréquence p de l'allèle C au bout de 40 ans (donc en 1990) dans le Dorset puis à Birmingham.
Dans les années 1950, la Grande-Bretagne adopta une législation anti-pollution (« the Clean Air Acts ») qui eut pour effet de réduire les émissions de suie et de SO2. Dans la période qui suivit, on observa une diminution de la fréquence de la forme mélanique, qui se poursuit depuis lors. Le graphe ci-dessous présente les observations effectuées dans une banlieue en voie de désindustrialisation.

8) Comparez la fréquence p de l'allèle C attendue à Bimingham en 1990 si la région était restée aussi industrielle (d'après le modèle mathématique) et la fréquence réellement observée à la même période dans une banlieue en voie de désindustrialisation. Justifiez qu'une innovation puisse se révéler tantôt favorable, tantôt défavorable selon les contraintes sélectives.
|
Remarque : l'expérience de KETTLEWELL, bien que concluante, est criticable sur la méthode. En effet, dans la nature, les phalènes ne s'exposent pas sur les troncs à leurs prédateurs mais se cachent sous les feuilles...Cette maladresse, non remarquée à l'époque, est utilisée par les créationnistes pour réfuter la théorie synthétique de l'évolution. Pourtant les exemples à l'appui de la sélection naturelle ne manquent pas (cf cours!). Alors pourquoi prendre l'exemple de la phalène ?...Parce que c'est une expérience historique très démonstrative que l'on trouve dans tous les manuels d'SVT et qui peut tomber au bac!
Pb : nous venons de voir que la fréquence d'un allèle qui confère un avantage sélectif augmente dans une population au fil du temps. Comment évoluerait la fréquence d'un allèle qui ne confère ni un avantage ni un désavantage sélectif ?
Hypothèse :
|
II- Une approche expérimentale de la dérive génétique
40 boules de différentes couleurs représentant les différents allèles d'un gène sont enfermées par paires dans 20 sacs opaques représentant les 20 individus d'une population.
Il y a en tout 10 boules blanches, 10 boules vertes, 10 boules rouges et 10 boules bleues mais la répartition dans les sacs n'est pas connue.
Dites quelles sont les fréquences alléliques dans la population de départ.
Pour simuler l'action de la reproduction sexuée au sein de cette population, mettez de côté au hasard 4 sacs qui représentent les individus qui n'auront pas de descendants à la génération suivante. Formez 8 couples avec les 16 sacs restant.
Pour chacun des couples, effectuez un tirage au sort en prenant une boule dans chaque sac, notez le couple d'allèles possédé par ce premier "enfant" et remettez les boules dans leur sac d'origine.
Faites un deuxième tirage au sort en ne prenant que 6 couples au hasard parmi ceux qui avait déjà formé un "descendant", notez les génotypes des 6 enfants formés et remettez les boules en place.
Faites un troisième tirage au sort pour 4 couples pris au hasard parmis ceux qui avaient eu un second "enfant" puis pour 2 couples pris au hasard parmis ceux qui avaient eu un troisième enfant.
Vous disposez maintenant des génotypes de 20 individus issus de la première génération.
1) Calculez les fréquences alléliques dans cette population. Que constatez-vous ? Confrontez vos résultats à ceux des autres groupes.
|
2) Quelle relation pouvez-vous faire entre ce résultat et l'action de la sélection naturelle ?
|
Lien vers un logiciel de simulation permettant de tester l'évolution de la fréquence d'un allèle sélectivement neutre sur 100 générations :
http://darwin.eeb.uconn.edu/simulations/selection-drift.html